Part 1, Introduction
Jeremy R. Young, Palaeontology Dept., NHM, Cromwell Road, London, SW7 5BD, UK &
Paul R. Bown, Dept. of Geological Sciences, UCL, Gower Street, London, WC1E 6BT, UK
The original version of this ms was printed in the Journal of Nannoplankton Research, issue 19/1
We have recently been working on a new version of A Stratigraphical Index of Calcareous Nannofossils (Bown, 1998). During this work we needed to address the question of the classification of nannofossils, and attempted a comprehensive generic-level review. We had originally intended to publish this in the book but space constraints prevented this. In addition, since one of the problems of classification is a lack of consensus, it seemed sensible to proceed on a broader basis. So, following consultation with various INA committee members, we decided to publish a working draft classification in the Journal of Nannoplankton Research in good time for subsequent discussion at the Puerto Rico INA Conference, at which we will host a workshop during which the following papers will be thoroughly discussed. Eventually, it is our intention to publish a final version of these papers, incorporating the results of these workshop discussions, as a handbook which will be distributed to all members of the INA. We have set out the classification in three separate papers. First, a discussion of the higher-level approach adopted and its relationship to previous classifications, followed by separate sections on Mesozoic and Cenozoic nannofossils. The Mesozoic/Cenozoic divide follows the precedent of Perch-Nielsen (1985a, b), and for much the same reasons: most major groups are predominantly confined to one era or other, so that grouping Mesozoic and Cenozoic nannofossils together in taxonomic sequence results in awkward alternations between the two eras. An overview of the classification is given in Figure 1
The starting point for this overview was the comprehensive review and classification of nannofossils of Perch-Nielsen (1985a, b), complimented by, in particular, the Jurassic nannofossil classification of Bown (1987, 1996) and the classification of extant haptophytes by Jordan and co-workers (Kleijne, 1993; Green & Jordan, 1994; Jordan & Kleijne, 1994; Jordan & Green, 1994; Jordan et al., 1995). Important earlier sources for these classifications include Parke & Dixon (1976), Hay (1977), Okada & McIntyre (1977) and Tappan (1980). In order to provide a comprehensive synthesis, we paid special attention to taxonomic papers published since 1985, as catalogued in the INA Bibliography and Index of Taxa. For the Tertiary, the reviews of Aubry (1984 et seq.) were invaluable sources, although they are based on a slightly different taxonomic philosophy.
Top-down subdivisions vs bottom-up
groupings
![]()
Two main approaches can be identified in the classification of calcareous nannofossils. The top-down subdivision approach has attempted to classify the range of nannofossils into well-defined subdivisions using clear-cut criteria based on major aspects of shape or structure. For instance, Deflandre (1950) divided all nannofossils into ortholiths and helioliths based on whether they showed radial calcite orientations (helioliths) or were formed of single calcite crystals (ortholiths, including the constituent segments of pentaliths). The bottom-up grouping approach, by contrast, has been based on organisation of species into genera and of genera into families based on multiple similarities of morphology, existence of intergradational morphologies and stratigraphic distribution. The latter, bottom-up, approach is, we believe, more productive and leads to a more natural classification, and it is the predominant method applied in Perch-Nielsen (1985a, b). This forms the basis for our formal classification into genera, families and orders. The top-down subdivision approach is, however, valuable for organising taxa of uncertain inter-relationship and so we apply a system of this type to arrange the higher taxa we recognise in a logical sequence, and in some cases to provide arbitrary subdivisions of particular taxa.
Incertae sedis and informal categories
![]()
A key objective of classification is to systematise our existing knowledge. Conversely, an obvious pitfall of classification is to provide, via spurious pigeon-holing, the impression of knowledge where none exists. To avoid this we have attempted to discipline ourselves and only to include genera within families (or families within orders) where there is good positive evidence for doing so. As a consequence, the classification is not comprehensive - many genera are left as of uncertain family/order, and many families are not assigned to an order. We do not apologise for this, since it is a simple reflection of the limits of current knowledge.
Use of orders as well as families ![]()
The only higher taxonomic category which has been universally applied is family and we continue to use families as the main level of classification. However, in revising the classification, we considered that exclusive use of families was unduly restrictive and that, with around 50 families, important relationships would be obscured.
We decided to use orders, rather than to subdivide families, in order to retain as many taxa as possible at their current level, the alternative being to reclassify many well-established families as subfamilies. Also, orders have previously often been used in nannofossil classifications, notably by Hay (1977), Tappan (1980) and Bown (1987). The principle potential problem here is discordance with use of orders in the classification of modern haptophytes. Classifications of modern haptophytes have, following Parke & Green (in Parke & Dixon, 1976), usually recognised four orders: Coccosphaerales (coccolith-bearing), Prymnesiales (typical non-coccolith-bearing haptophytes, haptonema well developed), Isochrysidales (haptonema diminutive, including some coccolith-bearing genera), and Pavlovales (with the flagella of unequal lengths, the longer flagellum with hairs and scales, haptonema diminutive). Recent work on molecular genetics, cytology and biochemistry, have unequivocally established the distinctive nature of the Pavlovales and they are now separated as a discrete class or subclass (Green & Jordan, 1994). The subdivision of the remaining haptophytes into the other three orders has not, however, been supported by subsequent studies, with the result that Green & Jordan (1994) recommended subsuming all these orders into the single subclass Prymnesiophycidae, order Prymnesiales. Given this abandonment of the traditional ordinal-level classification of extant haptophytes there is no obvious problem in introducing a new ordinal-level classification based in the first place on coccolith morphology, although obviously future research on aspects of haptophyte biology is liable to result at least in modifications of this classification. Nonetheless, this is not an area where we are especially dogmatic, and debate on use of particular levels of classification would be welcome.
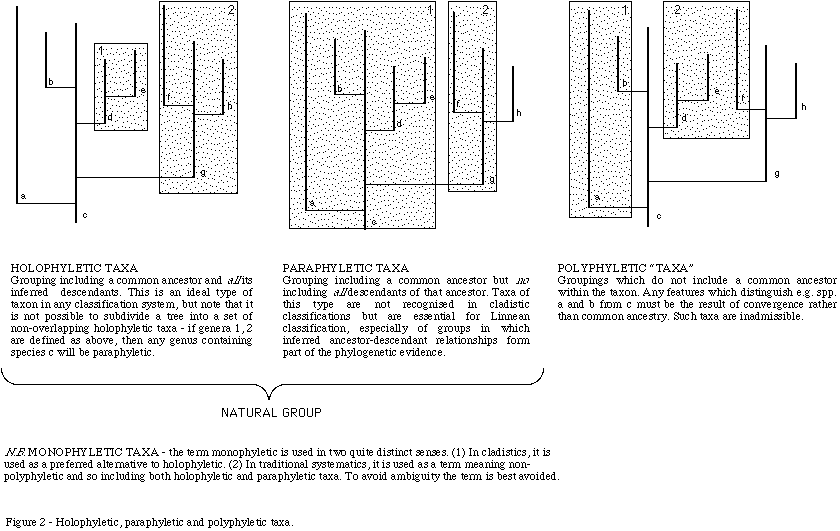
Paraphyletic and polyphyletic taxa ![]()
We have not attempted to avoid paraphyletic taxa, i.e. taxa which give rise to other taxa of the same, or higher, level (Figure 2). For instance, according to our current understanding of coccolithophore phylogeny, the family Zygodiscaceae is paraphyletic since it is inferred to have given rise to the families Pontosphaeraceae and Helicosphaeraceae. Similarly, the genus Lophodolithusis paraphyletic if we accept the interpretation that it gave rise to the genus Helicosphaera and family Helicosphaeraceae. This is manifestly an unavoidable problem in nannofossil systematics since we have abundant evidence of ancestor-descendant relationships. To give an extreme example, the genera Praeprinsius, Prinsius, Toweius, Reticulofenestra, Gephyrocapsa, and Emiliania are widely inferred to constitute an evolutionary lineage (e.g. Romein, 1979; Gallagher, 1989) and so all these, except Emiliania, are paraphyletic.
On the other hand, it is clearly essential to eliminate polyphyletic taxa, i.e. all species included in a higher taxon must have a common ancestor within that taxon (Figure 2). This is an absolute necessity of a biologically meaningful classification - if available evidence suggests that a taxon contains unrelated sets of species then it must be revised. Of course, we often do not know for certain whether a group of more or less similar species is directly related or simply a set of convergent homeomorphs. All taxonomic groupings may be regarded as hypotheses which may be tested through analysis of morphology and direct evolutionary data. Our general approach has been to avoid giving taxonomic names to any subdivisions which we feel are likely to be polyphyletic, for instance we have followed Kleijne (1991) in dividing the large number of extant holococcolith-bearing genera into two groups based on whether they show dimorphism or not. However, since this character is very likely to have evolved repeatedly, it does not form a suitable basis for grouping the genera into formal taxa.
Heterococcoliths,
holococcoliths and nannoliths
![]()
A special problem with nannofossil taxonomy is the uncertainty as to whether the structures we are examining are homologous. Indeed, we know that holococcoliths and heterococcoliths are not homologous, whilst we are uncertain as to the nature of nannoliths. This requires some discussion, and in addition there are separate taxonomic problems associated with each of these groupings.
This is the predominant group, usually defined as coccoliths formed of a limited number of complex and variably shaped crystal-units. The precise scope of the term heterococcolith is vague but there is no dispute that the term is based on typical extant coccoliths such as those of Coccolithus, Emiliania, Pleurochrysis and Syracosphaera (see also original description of Braarud et al., 1955). These all share the basic morphology of a rim formed of a radial array of complex crystal-units surrounding a central-area, with variable structures in the central-area. This basic morphology characterises numerous living and fossil groups and forms an essential part of most workers' concept of an heterococcolith. There is also evidence that this is a natural, non-polyphyletic, grouping: (1) studies of evolution in the group tend to indicate a pattern of common origin and subsequent divergence, with no obvious indications of polyphyly (e.g. Bown, 1987; Romein, 1979; Bown et al., 1992); (2) our ultrastructural research (Young & Bown, 1991; Young et al., 1992; unpublished data) suggests that a widespread, and probably universal, feature of heterococcoliths is V/R mode biomineralisation, i.e. biomineralisation which commences with nucleation of a proto-coccolith ring of alternating subvertical and subradial crystal-units, which subsequently develop to form the rim. This type of biomineralisation is characteristic of all the heterococcolith families recognised here except possibly the Rhabdosphaeraceae (only V-units definitely identified), Hymenomonadaceae and Papposphaeraceae. V/R mode biomineralisation is a distinctive and sophisticated type of calcification and it seems reasonable to assume that it is a reliable synapomorphy (shared derived character state), and that the heterococcoliths as a whole form a non-polyphyletic group.
Holococcoliths are plainly formed by a very different biomineralisation process to heterococcoliths. Nucleation must occur across the entire coccolith and the individual crystals retain simple euhedral morphologies. As a result, there is no clear distinction between the rim and central-area. Moreover, available evidence suggests that calcification occurs extra-cellularly (Rowson et al., 1986), rather than intracellularly. Finally, there are now several well-established cases of holococcoliths and heterococcoliths occurring on alternate phases of the life-cycle of single species. Billard (1994) synthesises this evidence and suggests that it reflects a haplodiplontic life-cycle, with holococcoliths consistently occurring in the haploid phase. This rather suggests that biomineralisation may have evolved independently in the haploid and diploid phases, and that holococcoliths and heterococcoliths are not meaningfully homologous. Possibly we should consider holococcolith and heterococcolith taxonomy as independent, much as spore and whole-plant taxonomy are. Figure 3 shows how the rather disparate set of observations on coccolithophorid life cycles can be rationalised into a simple phylogeny.

Holococcolith biomineralisation is distinctive and sophisticated. Crystallite faces are perfectly aligned across large zones, implying precise control of both a- and c-axis orientation, and the uniformity of crystallite size implies similarly precise control of nucleation spacing. Like V/R mode biomineralisation in heterococcoliths, it is unlikely that the holococcolith biomineralisation mode has evolved repeatedly, rather we can regard holococcolith formation as evidence for common descent of all taxa in which it is developed. Plainly though, both holococcolith formation and heterococcolith formation may have been lost repeatedly during the evolutionary history of haptophytes. So there is no reason for assuming that all holococcoliths will have heterococcolith-bearing phases, or that all species which are known to have both holo- and heterococcolith-bearing phases will be particularly closely related. Future research will certainly help elucidate relationships. In the interim, it is entirely logical for all holococcolith bearing taxa, without known heterococcolith phases, to be classified together, in the family Calyptrosphaeraceae.
Unlike heterococcoliths, holococcolith biomineralisation does not provide obvious characters for reliably subdividing the group. Homology is difficult to establish since the basic structure appears to consist of a simple mass of crystallites, gross morphology appears to be of limited value, and there are serious problems due to dimorphism. So at present, subclassification of the Calyptrosphaeraceae is confined to a rather unsatisfactory, and probably partially artificial, generic classification. This applies particularly to extant holococcoliths for which crystallographic orientations are poorly known. In the larger fossil holococcoliths, subdivision into blocks with predictable crystallographic orientations appears to provide a reasonable basis for classification, but original morphology and possible dimorphism cause separate problems.
When Braarud et al. (1955) defined the terms heterococcolith and holococcolith, they explicitly described the pentaliths as a third category of similar status. This was a logical proposal since pentaliths show different structures to any heterococcoliths and it is reasonable to infer they have formed by a significantly different growth/biomineralisation process, and therefore are not directly homologous. The same applies to numerous other groups, particularly amongst the extinct nannofossils, e.g. sphenoliths, nannoconids and discoasters. The term nannoliths has been widely used as a collective term for this group (e.g. Perch-Nielsen, 1985a, b; Bown, 1987; Aubry, 1984 et seq.; Young et al., in press). The most common definition is calcareous nannofossils of uncertain affinity, but probably related to coccolithophores, i.e. formed by haptophytes.
Although nannoliths are highly variable, several features are repeatedly shown: (1) most nannoliths have morphologies which are at least suggestive of having been part of a composite exoskeleton; (2) most nannoliths are formed of morphologically-complex crystal-units analogous to those of heterococcoliths, although in some cases with evidence of additional ultrastructure (e.g. laminae within pentalith crystal-units); (3) for all known nannoliths there are consistent relationships between morphology and crystallographic orientation; (4) most nannoliths definitely show consistent handedness or chirality (deviation from radial symmetry), e.g. nannoconids always show clockwise imbrication, Braarudosphaera sutures are always offset clockwise from the median point of the pentalith edge; (5) nannoliths typically have biogeographic patterns suggesting planktonic origin and as a group are only known from the Triassic to Recent.
These features all suggest close affinities with coccolithophores. Moreover, haptophytes are the only plankton group known to produce calcareous scales. Dinoflagellates also calcify but they form calcispheres rather than discrete scales. Various other phytoplankton produce scales, e.g. Prasinophytes and Chrysophytes, but these are either unmineralised or siliceous. Hence, an assumption that nannoliths are produced by haptophytes has seemed reasonable to phycologists and has been supported by independent evidence in several cases, e.g. association of ceratoliths and coccoliths (Norris, 1965; Alcober & Jordan, 1997), observation of haptonema on Polycrater (Thomsen et al., 1994), LM observations of typical haptophyte cell-features for Braarudosphaera(J. Green, pers. comm., 1996).
Nannoliths as a whole are, however, almost certainly polyphyletic, probably including: (1) modified heterococcoliths (?Florisphaera , ?polycycloliths, ?helioliths, ?fasciculiths, ?sphenoliths); (2) modified holococcoliths (?ceratoliths, ?Lithostromationaceae); (3) and nannoliths formed by entirely different biomineralisation mechanisms (?pentaliths, ?Polycrater, ?nannoconids). So they cannot be regarded as a formal taxonomic group, even though it is extremely useful to distinguish them from the more rigorously definable holo- and heterococcoliths. Further research will probably allow some to be reclassified as aberrant heterococcoliths or holococcoliths, whilst others may prove to be discrete groups.
The haptophytes were traditionally included in the phylum Chrysophyta along with a range of other golden-brown, scale-bearing algae. However, Parke et al. (1955) recognised the haptonema as a unique structure, and following this they were recognised as a distinct class by Christensen (1962) and division by Hibberd (1972). Rather unfortunately, Hibberd (1976) argued that the name Haptophyta was unsuitable since it was not based on a generic root. He proposed the alternative name Prymnesiophyta, and this name was widely used in the literature until recently. However, as noted by Green & Jordan (1994), under the International Code of Botanical Nomenclature, division-level names do not need to be typified and the name Haptophyta has priority. Following from this, the common term haptophyte is preferable to prymnesiophyte; it is also a less forbidding term so we expect it to rapidly regain universal currency.
Notwithstanding this nomenclatural confusion, the separation of the haptophytes on the grounds of flagellar character has been well supported by much other evidence from cytology, biochemistry and molecular genetics. There is no dispute that they form a discrete algal protist group separable at division/phylum level. Chloroplast colour, however, reflects rather fundamental aspects of pigment biochemistry, e.g. green algae and land plants all have chlorophyll a and b. By contrast, golden-brown algae, including haptophytes, have chlorophyll a and c. This unites them with a range of other non-green algae, including the diatoms and silicoflagellates. Another common feature of this group is that the chloroplasts are surrounded by four membranes, which is interpreted as the result of a complex endosymbiosis history. Finally, most of these groups have two flagella, one short and smooth, the other long and bearing tubular hairs (heterokont). On the basis of this set of shared characters, Cavalier-Smith (1981, 1994) has proposed removing the group from the Protista to form a separate kingdom, Chromista, also including various protozoa which show the same heterokont flagellation. The haptophytes, as noted above, have two smooth flagella, and so do not show heterokont flagellation, hence their position in this group is somewhat uncertain, but the general interpretation is that they are a primitive group that diverged prior to the evolution of heterokont flagellation (e.g. Cavalier-Smith, 1994).
Within the haptophytes, only the single class Prymnesiophyceae is now recognised, divided into two subclasses, the Prymnesiophycidae and Pavlovophycidae (see Green & Jordan, 1994). All coccolithophores belong to the Prymnesiophycidae, and so to:
Kingdom CHROMISTA Cavalier-Smith, 1981
Division (Phylum) HAPTOPHYTA Hibberd ex Cavalier-Smith, 1986
Class PRYMNESIOPHYCEAE Hibberd, 1976
Subclass PRYMNESIOPHYCIDAE Cavalier-Smith, 1986
The same format has been adopted for both of the following papers. For each genus, the original authorship is given, followed by important synonyms in brackets (). There are then short notes outlining the basis for distinguishing the genus. Terminology used in these descriptions follows Young et al. (in press). In some places we have included genera which are arguably redundant, and these are placed in square brackets []. Uncertain assignments are indicted by a question mark.
The formal systematic classification is supplemented, as seems useful, by informal groupings. These are numbered and given in bold. In most cases, these subdivisions are arbitrary and are likely to prove artificial/polyphyletic, hence they are not proposed as the bases for future formal classification.
Acknowledgements: Thanks to Katharina von Salis, Jackie Burnett, Liam Gallagher, Matt Hampton and Kevin Cooper for comments and discussion at various stages of this work, and to Jackie Burnett and Bohumil Hamrsmid for their fine work in converting our text to a uniform publishable format.
References Constraints of space meant it was not practical to provide a complete bibliography of taxonomic references here. The references can, however, be found in Perch-Nielsen (1985a, b), the INA Bibliographies, or on Wuchang Wei's online nannofossil bibliography. References cited in this introduction or in the comments sections of the maintext are listed on a separeate page.