8th International Nannoplankton Association Conference
ABSTRACTS
[Abstracts] [A] [B] [C] [D] [E] [F] [G] [H] [I] [J] [K] [L] [M] [N] [O] [P] [Q] [R] [S] [T] [U] [V] [W] [X] [Y] [Z]
Hanno Kinkel, Karl-Heinz Baumann, Markus Geisen, Ian Probert, Heather Stoll, Jeremy Young, Patrizia Ziveri:
Size matters: the influence of coccolith size and growth variation on reconstructing past carbon cycles
(Talk)
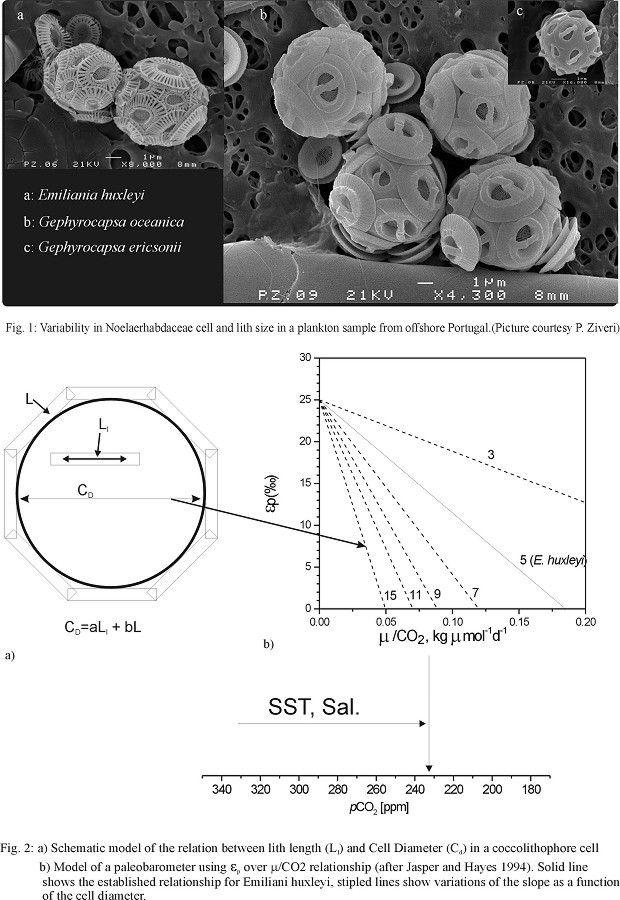
Coccolith morphometry has been a standard tool in nannoplankton research. In general, it is used to separate similar forms into species, subspecies or morphotypes, based on simple measurements, such as length and width, and other morphometric features, such as pore-size or rim-width, are considered as well. Recently, Young & Ziveri (2000) proposed a method to estimate carbonate masses of single coccoliths using morphometric measurements and a geometric model. Thus, it is possible to calculate species-dependent carbonate fluxes of coccoliths, which have changed considerably during time.
Coccolith calcification is considered to contribute significantly to global carbon cycles, both by adding CO2 during calcification, which is a short-term effect, and on the other hand by storing huge amounts of carbon in the sedimentary archives, where they reside for long times.
Long-chain alkenones, biomarkers synthesised almost exclusively by species of the Family Noelaerhabdaceae, provide useful proxies for palaeoceanographers. Their unsaturation ratio is linearly correlated with sea-surface temperature (SST: Prahl & Wakeham, 1987), whereas their stable carbon isotope composition is thought to provide the most useful proxy for reconstructing past pCO2 values of surface-waters (Jasper & Hayes, 1994). The isotopic fractionation ep of algal biomass is strongly influenced by the surface-area to volume ratio of the algae (Popp et al., 1998). Since coccolithophores calcify intracellularly, their cell-volume is proportional to the length of their liths, and thus coccolith morphometrics provide the only possibility to estimate cell geometry of extinct phytoplankton species. Fortunately, most morphometric information is available for the group of coccolithophores that are producing alkenones, the Noelaerhabdaceae (Bollmann, 1997; Samtleben, 1980; Young, 1990). Thus, a large dataset on size variations already exists which, upto now, has been ignored in geochemical studies. In combination with other geochemical proxies derived from coccolith carbonate (Sr/Ca, 13Ccarb), we try to reconstruct size-related changes in coccolithophore carbonate budgets and the implications for the global carbon cycle.
 |
References
Bollmann, J. 1997. Morphology and biogeography of Gephyrocapsa coccoliths in Holocene sediments. Marine Micropaleontology, 29: 319-350.
Jasper, J.P. & Hayes, J.M. 1994. Reconstruction of paleoceanic pCO2 levels from carbon isotopic compositions of sedimentary biogenic compounds. Springer, Berlin: 323-341.
Popp, B.N., Laws, E.A., Bidigare, R.R., Dore, J.E., Hanson, K.L. & Wakeham, S.G. 1998. Effect of phytoplankton cell geometry on carbon isotopic fractionation. Geochimica et Cosmochimica Acta, 62: 69-77.
Prahl, F.G. & Wakeham, S.G. 1987. Calibration of unsaturation patterns in long-chain ketone compositions for paleotemperature assessment. Nature, 330: 367-369.
Samtleben, C. 1980. Die Evolution der Coccolithophoriden-Gattung Gephyrocapsa nach Befunden im Atlantik. Paläont. Z., 54: 91-127.
Young, J.R. 1990. Size variation in Neogene Reticulofenestra coccoliths from Indian Ocean DSDP cores. Micropaleontology, 9: 71-86.
Young, J.R. & Ziveri, P. 2000. Calculation of coccolith volume and its use in calibration of carbonate flux estimates. Deep-Sea Research.
[INA8]
[Registration and Accomodation
Form]
[First Circular and Pre-Registration]
[Second Circular]
[Division of Micropalaeontology] [Department of Geosciences] [Bremen University]
Copyright © 2000, most recent revision July 24, 2000
Tania Hildebrand-Habel (hiha@micropal.uni-bremen.de)